ЭКСПРЕССИЯ ГЕНОВ И МЕЖПОЛУШАРНАЯ АСИММЕТРИЯ
Научная статья: «ЭПИГЕНЕТИЧЕСКАЯ РЕГУЛЯЦИЯ ЛАТЕРАЛИЗИРОВАННОЙ ПОЗВОНОЧНОЙ ЭКСПРЕССИИ ГЕНОВ ПЛОДА ОБУСЛАВЛИВАЕТ МЕЖПОЛУШАРНЫЕ АСИММЕТРИИ (ЭКСПРЕССИЯ ГЕНОВ И МЕЖПОЛУШАРНАЯ АСИММЕТРИЯ)»
Себастиан Окленбург / Юдит Шмиц / Захра Моинфар / Дирк Мозер / Рена Клозе / Штефани Лор / Георг Кунц / Мартин Тегентофф / Педро Фаустман / Клайд Франкс / Йорг Т Эпплен / Роберт Кумста / Онур Гюнтюркюн
01.02.2017
Аннотация
Латерализация представляет собой основополагающий принцип организации нервной системы, но ее молекулярные детерминанты по большей мере неизвестны у людей. Предполагалось, что асимметричная экспрессия генов в плодной коре надпочечника являлась молекулярной основой праворукости или леворукости. Однако, для человеческих плодов характерна значительная асимметрия в движении руками еще до того, как двигательная область коры головного мозга функционально связывается со спинным мозгом, делая более вероятным предположение, что <b>асимметрии позвоночной экспрессии генов формируют молекулярную основу леворукости или праворукости</b>. Мы проанализировали экспрессию матричной РНК по всему геному и метилирование ДНК в цервикальном и переднем грудном сегменте спинного мозга у пяти человеческих плодов и продемонстрировали асимметрию экспрессии генов, зависящую от развития. Данная асимметрия экспрессии генов эпигенетически управляется асимметрией экспрессии микроРНК в сигнальном пути TGF-β и латеризированным метилированием CpG – островков. На основе наших результатов мы предполагаем, что молекулярный механизм эпигенетической регуляции внутри спинного мозга является отправной точкой для леворукости или праворукости, предполагая основное изменение в нашем понимании онтогенеза межполушарной асимметрии у людей.
Введение
По сравнению с почти бесконечной многокомпонентностью мыслительного процесса и поведения позвоночных, количество генов, оказывающих влияние на развитие центральной нервной системы поразительно небольшое (Кадаккужа и Путанвееттил, 2013). Поэтому, понимание молекулярного механизма, обусловленного эпигенетикой архитектуры центральной нервной системы позвоночных стало центральным вопросом неврологии (Кундакович и Шампан, 2015).
Нашим основополагающим принципом организации головного мозга является латерализация, т.е. структурное или функциональное различие между левым и правым полушариями головного мозга (Корбаллис, 2014). Латерализация представляет собой консервативную характеристику всех позвоночных (Штрекенс и др.,2013; Окленбург и др., 2013a; Бисацца и др., 1998; Роджерс и др., 2012; Версаче и Валлортигара, 2015) и в последних исследованиях активно предполагается, что она также присутствует и у беспозвоночных (Фраснелли и др., 2012). Данная повсеместность поведенческой и мозговой латерализации решительно поддерживает идею о том, что латерализованная организация центральной нервной системы предлагает эволюционное преимущество (Валлортигара и Роджерс, 2005).
К предположениям о том, почему латерализованный мозг мог бы повысить хорошее физическое состояние организма, относятся предотвращение ненужной дупликации активности нейронов в обоих полушариях, более быстрые нейрональные процессы, благодаря отсутствию медленного трансфера информации посредством мозолистого тела между полушариями и более хорошая координация одностороннего поведения в стаях или других социальных группах животных (Валлортигара и Роджерс, 2005; Корбаллис, 2009). У людей межполушарная асимметрия проявляется почти во всех основных когнитивных системах (Окленбург и др., 2014а), включая язык (Фридеричи, 2011; Сепета и др., 2016), память (Джамматтеи и Арндт, 2012; Тат и Ацума, 2016; Хабиб и др., 2003), внимание (Фаласка и др., 2015; Дрюкер и др., 2013), эмоциональные процессы (Дамарее и др., 2005), восприятие (Де Винтер и др., 2015), кратковременная память (Лангель и др., 2014; Нагель и др., 2013) и управляющие функции (Окленбург и др., 2011а, 2012; Шток и Бксте, 2014).
Несомненно, самым исследуемым проявлением латерализации у людей является леворукость и праворукость (Корбаллис, 2014). Важно отметить, что леворукость и праворукость относятся к латерализированной организации когнитивных систем в человеческом мозге (Окленбург и др., 2014b; Фрессле и др., 2016). Например, левополушарное языковое доминирование установлено у 96% праворуких субъектов, и лишь у 73% леворуких субъектов (Кнехт и др., 2000). Актуальность леворукости и праворукости недавно была подчеркнута Виллемсом и др. (Виллемс и др., 2014), который утверждает, что это является одним из самых важных факторов, влияющих на индивидуальную организацию головного мозга, и что явное включение леворуких людей в экспериментальные исследования значительно улучшает наше понимание языка, двигательного поведения и зрительной обработки.
Леворукость и праворукость не только служат в качестве поведенческого посредника для индивидуальной организации головного мозга, они также интересны в плане клинической перспективы: большое количество психоневрологических расстройств и болезней, связанных с нарушением развития, таких как расстройства аутистического спектра (Колби и Паркисон, 1977; Форрестер и др., 2014; Преслар и др., 2014; Райсстад и Педерсен, 2016), депрессии (Денни, 2009; Элиас и др., 2001; Лог и др., 2015), биполярные аффективные расстройства (ван Дайк и др., 2012; Новаковская и др., 2008), тревожные расстройства (Лог и др., 2015; Хикс и Пеллегрини, 1978; Орме, 1970; Райт и Харди, 2012; Харди и др., 2016; Лайл и др., 2013), шизофрения (Хирнштейн и Хугдаль, 2014; Драгович и Хаммонд, 2005; Зоммер и др., 2001) или алкоголизм (Денни, 2011; Мандаль и др., 2000; Сперлинг и др., 2000) ассоциировались с леворукостью или смешанным доминированием рук. Поэтому, понимание онтогенеза леворукости или праворукости, а также межполушарной асимметрии в целом может привести к потенциально важным выводам относительно патогенеза данных расстройств.
Однако, несмотря на их важность для многих аспектов организации головного мозга, онтогенетические основы асимметрии головного мозга все еще далеки от полного понимания. Изначально предлагались теории единичных генов для объяснения появления леворукости и праворукости в качестве функции одного гена с двумя аллелями (Аннетт, 1998; МакМанус, 1985). Однако, в процессе недавно проведенных общегеномных исследований ассоциации не удалось обнаружить ни одного общегеномного значительного однонуклеотидного полиморфизма, что опровергает теорию единичного гена (Армур и др., 2014; Эрикссон и др., 2010). При проведении исследований кандидатных генов обнаружилось множество генов, которые отображали ассоциацию с леворукостью, в том числе трансмембранный нейрональный белок с лейцин-богатыми повторами 1 (LRRTM1) (Франкс и др., 2007), пропротеинконвертаза 6-го субтилизин-кексинового типа 6 (PCSK6) (Скерри и др., 2011; Арнинг и др., 2013; Брандлер и др., 2013) и ген андрогенового рецептора (AR)(Арнинг и др., 2015; Хэмпсон и Санкар, 2012; Медланд и др., 2005). Однако, данные гены объясняют только малую долю вариаций в данных о леворукости. Более того, в большом количестве исследований предполагалось, что только примерно четверть вариаций леворукости или праворукости связаны с генетическими изменениями, в то время как оставшиеся 75% вариаций объясняются факторами окружающей среды, несвязанными с генами (Медланд и др., 2006; Вуоксима и др., 2009). Данные результаты подчеркивают важность интеграции как генетических изменений, так и эпигенетических процессов, модулирующих экспрессию генов, в процессе исследования онтогенеза межполушарной асимметрии (Гешвинд и Миллер, 2001).
Предполагалось, что асимметричная экспрессия генов является молекулярной основой различия при выборе руки, которая будет использоваться: Сун и др. (Сун и др., 2005) сравнивали уровни экспрессии генов в правом и левом персильвиарных отделах коры головного мозга человеческого плода. На 12 неделе беременности авторы идентифицировали 27 постоянно асимметричных экспрессированных генов, которые, по большей мере, несли ответственность за регуляцию экспрессии генов, сигнальную трансдукцию и развитие коры. Одним из постоянно асимметричных экспрессированных генов был рекомбинатный белок человека 4 (LMO4). Дальнейшие исследования показали, что одностороннее отклонение выражения Lmo4 при воздействии на эмбрион мышей замедляет нейрогенез в одном полушарии, что ведет к образованию асимметричной функциональной области, производству нейронов и аксональных отростков, а также изменяет предпочтение при выборе руки, которая будет использоваться (Ли и др., 2013). Анализ экспрессии генов в головном мозге взрослого человека дал меньше ясных результатов, в процессе двух независимых исследований не удалось найти различия в экспрессии генов между аналогичными участками полушарий головного мозга (Гаврилич и др., 2012; Плетикос и др., 2104). В недавно проведенном исследовании Карлебах и Франкс повторно проанализировали оба массива данных и продемонстрировали, что едва уловимая латерализация на уровне единичного гена преобразуется в более выраженную асимметрию на уровне функциональных групп генной онтологии (GO). Авторы установили, что наборы латерализованных генов необходимо связывать с электрофизиологией нейронов, синаптической передачей, развитием нервной системы и активностью глутаматных рецепторов (Карлебах и Франус, 2015).
Однако, недавно проведенные исследования указывают на то, что кортикальная ткань может и не представлять собой оптимальный выбор для исследования отношения асимметрии экспрессии генов и поведенческой асимметрии. Онтогенетически, леворукость или праворукость закладывается на раннем этапе развития, с момента появления скоординированных движений руками на 8 неделе после зачатия (ПЗ), т.е. при гестационном возрасте в 10 недель, когда 85% плодов демонстрируют больше движений правой рукой, чем левой. (Хеппер и др., 1998; Врайс и др., 1985). Исследование феномена сосания пальца у 274 плодов показало, что на 13 неделе после зачатия 90% предпочитают сосать большой палец правой руки, в то время как только 10% сосут большой палец левой руки (Хеппер и др., 1900, 1991). Интересно отметить, что последующее исследование 75 младенцев показало, что предпочтение в сосании пальца определенно связано с последующей леворукостью или праворукостью: 60 детей, предпочитающие сосать большой палец правой руки, становились праворукими, в то время как из 15 детей, предпочитавших сосать большой палец левой руки, пять стали праворукими, а десять – леворукими (Хеппер и др., 2005). Важно отметить, что двигательная область коры головного мозга функционально еще не связана со спинным мозгом на данном этапе развития, так как отростки корково-спинномозгового пути не входят в переднюю часть спинного мозга до 15 недель после зачатия (тен Донкелар и др., 2004). Отсюда следует, что леворукость или праворукость вряд ли находятся под контролем головного мозга (Хеппер и др., 1991) и асимметричные движения руки должны возникать в результате спинномозговой активности генов. Поэтому, вероятнее всего, скорее асимметрия экспрессии спинномозговых генов, чем генов, относящихся к коре, служит молекулярной основой леворукости или праворукости.
На асимметричный уровень экспрессии генов скорее всего оказывают влияние эпигенетические изменения. Самым важным эпигенетическим механизмом является метилирование ДНК. Связывание метиловых групп (-CH3) с CpG сайтами или островками становятся причиной снижения или предотвращения транскрипции и, соответственно, экспрессии генов. Подтверждено, что метилирование ДНК вовлечено в развитие основных функций центральной нервной системы, таких как синаптическая функция, пластичность нейронов, обучение, память (Николова и Харири, 2015; Дей и др., 2015; Рот, 2012). Показано, что на метилирование ДНК в особенности влияют внутриутробные факторы, вызывающие стресс (Туреки и Мини, 2016; Вайзерман, 2015), которые, в частности, интересны в контексте онтогенеза леворукости или праворукости. Более того, проведенные недавно исследования продемонстрировали, что метилирование играет роль в онтогенезе леворукости или праворукости: уровень метилирования в CpG блоке в промоторной области LRRTM1 связывают с атипичным выбором доминирующей руки (Лич и лр., 2014).
Посттранскрипционно, далее экспрессия генов регулируется посредством микроРНК (miRNAs), которые состоят из малых некодирующих РНК, длиной в 21-25 нуклеотидов. У людей и прочих млекопитающих микроРНК изначально становится причиной дестабилизации целевой мРНК, вместо снижения трансляции (Гуо и др., 2010). Это также релевантно для межполушарной асимметрии , так как нейтральная асимметрия у нематоды Caenorhabditis elegans контролируется при помощи различных миРНК (Алгадах и др., 2013; Кочелла и Хоберт, 2012., Джонстон и Хоберт, 2003; Хси и др., 2012).
Резкие изменения в пространственно-временных профилях экспрессии являются ключевой чертой человеческого эмбриогенеза (Йи и др., 2010), а образование функциональной асимметрии в позвоночных, как показано, сильно зависит от критических периодов на этапе раннего развития (Ле Гранд и др., 2003; Заппиа и Роджерс, 1983). Для исследования молекулярных детерминантов поведенческой асимметрии у человека мы проанализировали асимметрию в образцах общегеномной экспрессии мРНК, экспрессии микроРНК и метилирования ДНК в тканях спинного мозга человеческого плода. Важно отметить, что мы особо хотели исследовать сегменты спинного мозга, которые иннервируют руки и кисти. В то время как ростральные шейные сегменты (С2 – С5) иннервируют голову, шею и плечевую зону, последующие сегменты иннервируют напрямую руки и кисти, при этом С6 иннервирует большой палец, С7 – средний палец, а С8 – мизинец. T1 иннервирует среднюю сторону локтевой ямки (Майнард и др., 1997). По результатам исследования начального этапа лево-правой асимметрии в скоординированных движениях руками (Хеппер и др., 1998; де Врайс и др., 1985), мы заострили внимание на образцах ткани плода, полученных с 8 по 12 неделю после зачатия.
Мы предположили, что асимметрия экспрессии генов между левой и правой половиной спинного мозга начинает проявляться на 8 неделе после зачатия, так как с этого момента появляются скоординированные асимметричные движения руками. На основе результатов о роли негенетических факторов воздействия на развитие леворукости или праворукости мы также допустили явную модуляцию этой асимметрии экспрессии мРНК путем метилирования ДНК и экспрессии асимметричной микроРНК.
Результаты. Экспрессия генов
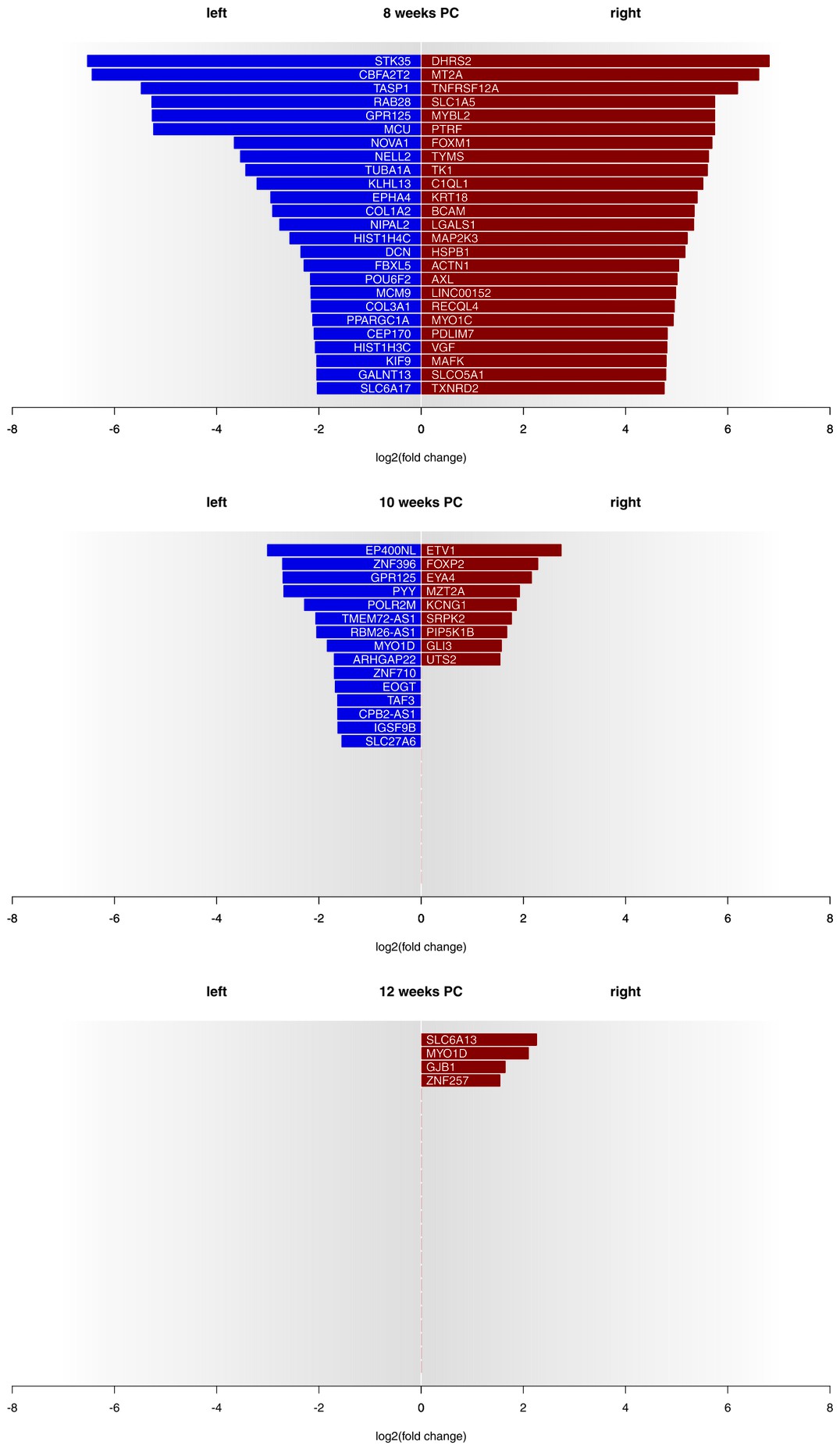
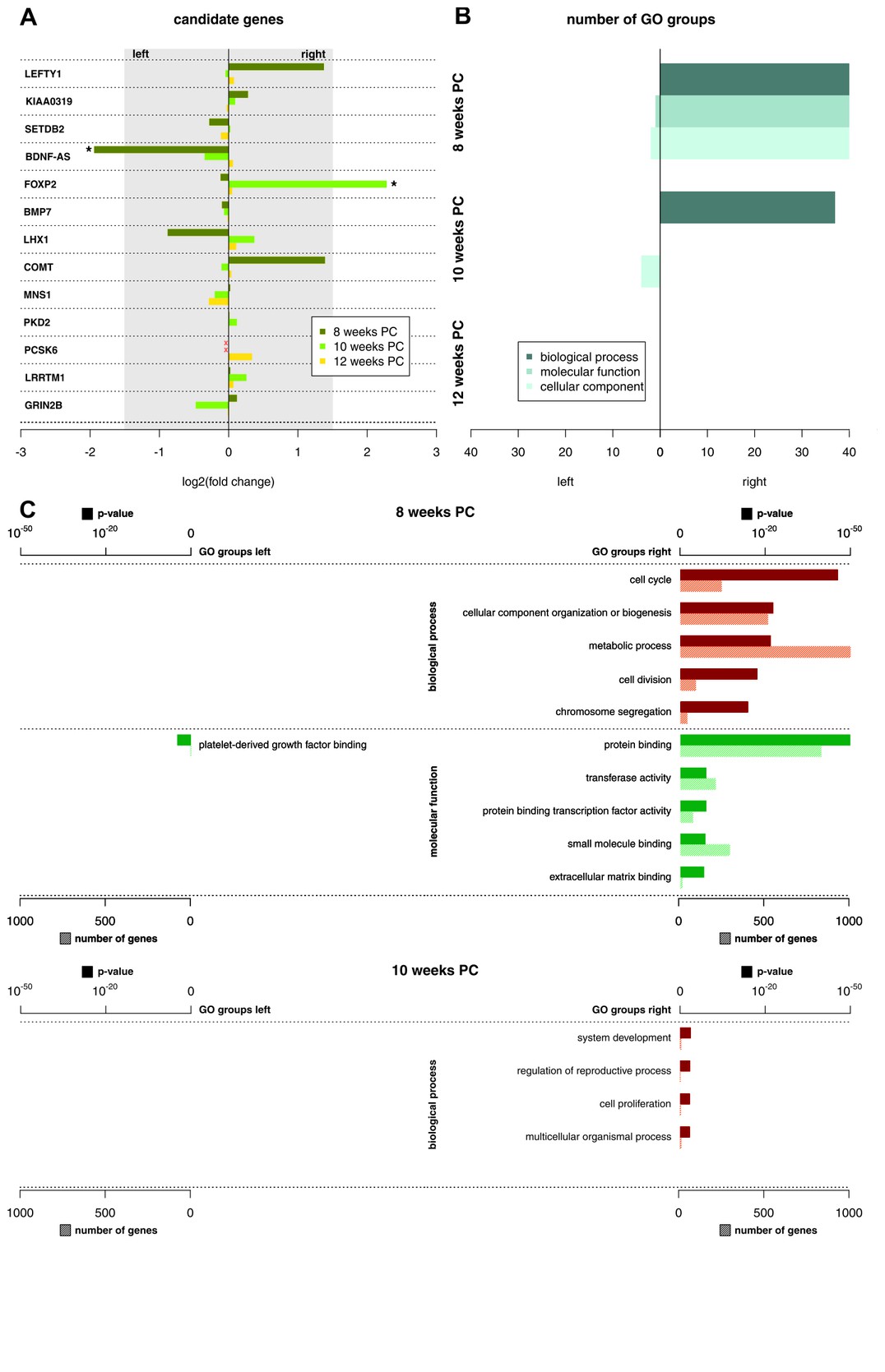
Асимметрия в экспресии мРНК в сегментах спинного мозга от С2 до Т2 были обнаружены на всех трех стадиях развития, с явно выраженными крупными различиями на 8 неделе после зачатия. На 8 неделе после зачатия 1690 транскриптов (3,29%) демонстрировали лево-правое различие при экспрессии генов с log2(кратность изменения) > 1.5. Тот факт, что 39 транскриптов демонстрировали более сильную левостороннюю экспрессию генов, в то время как 1651 транскрипт демонстрировали более сильную правостороннюю экспрессию генов, подчеркивает более сильную экспрессию генов с правой стороны на данной стадии развития. Количество асимметрически экспрессированных генов с log2(кратность изменения) > 1.5 уменьшилось только на 24 гена (0,05%) к 10 неделе после зачатия. Четыре гена (0.01%) продемонстрировали log2(кратность изменения) равную 1.5 или выше (см. Рисунок 1 для 25 основных асимметрически экспрессированных генов на стадию развития и Дополнительный файл 1G для индивидуальных образцов) на 12 неделе после зачатия, все эти результаты сигнализируют о более сильной экспрессии генов в правой половине спинного мозга. Среди кандидатных генов, связанных с развитием межполушарной асимметрии (см Рисунок 2А), forkhead box P2 (FOXP2)(Окленбург и др., 2013b) отображал правостороннюю асимметрию в спинном мозге на 10 неделе после зачатия. Мозговой нейротрофический фактор антимысловой РНК (BDNF–AS) (Маннс и др., 2008) был более экспрессирован в левой половине спинного мозга на 8 неделе после зачатия.
Обсуждение результатов
Межполушарные асимметрии в головном могзе и поведение представляют собой основные организационные принципы центральной нервной системы позвоночных, но их онтогенез еще плохо понятен (Окленбург и др., 2014b). Согласно общему мнению, как генетические, так и эпигенетические факторы играют роль (Окленбург и др., 2013с), но в то же время непонятно, какие молекулярные процессы обуславливают эпигенетическую модуляцию асимметрии экспресии генов, не выявлено потенциальное происхождение поведенческих асимметрий (Сун и др., 2005; Карлебах и Франкс, 2015). Для разъяснения данного вопроса мы провели анализ асимметрии в полногеномной экспресии мРНК, экспрессии микроРНК и метилировании ДНК в образцах тканей человеческого плода. Следует подчеркнуть, что мы заострили внимание на тканях спинного мозга, а не головного. Спустя восемь недель после зачатия, человеческие плоды демонстрировали явно выраженное латерализованное моторное поведение рук. Так как маловероятно, что такое поведение контролируется корой головного мозга (Хеппер и др., 1998; де Врайс и др., 1985; Хеппер и др., 2005; тен Донкелар и др., 2004), возникло предположение, что оно находится под контролем спинного мозга (Хеппер, 2013).
В соответствии с нашей гипотезой, на основании наших результатов можно предположить, что асимметрия экспрессии генов в сегментах спинного мозга, иннервирующих руки и кисти могла бы быть решающим моментом для онтогенеза функциональной асимметрии. Впервые мы указываем, что шейные и передние сегменты спинного мозга плода с левой и с правой стороны демонстрируют биологически релевантные различия в экспрессии генов. Важно отметить, что такая асимметрия экспрессии генов по большей мере обусловлена стадией развития. На 8 неделе после зачатия асимметрия экспрессии генов между двумя половинами спинного мозга проявляется очень четко, при этом 3,29% всех транскриптов демонстрировали биологически релевантные различия экспрессии генов с левой стороны, что гораздо больше относительно правой стороны, были вовлечены в многочисленные категории Генной онтологии, принимающие участие в неврологическом развитии. На 10 неделе после зачатия это количество существенно снизилось до 0,05% и так продолжалось на 12 неделе (0,01%). В то время как результаты для 10 и 12 недель большей частью сравнимы с результатами, полученными для асимметрии экспрессии генов в коре головного мозга плода (Сун и др., 2005; Карлебах и Франкс, 2015), данные относительно восьми недель после зачатия сигнализируют о значительном повышении, по сравнению с предыдущими отчетами, асимметрии экспрессии генов в тканях центральной нервной системы, что соответствует первому проявлению скоординированных движений руками.
Наряду с предположением о том, что факторы окружающей среды, несвязанные с генами, несут ответственность за более, чем 75% вариаций функциональной межполушарной асимметрии у человека (Медланд и др., 2009), мы смогли продемонстрировать, что большая часть таких асимметрий экспрессии генов управляется эпигенетическими процессами.
С одной стороны, мы смогли показать, что метилирование ДНК CpG островков демонстрирует значительную асимметрию, которая связана с асимметрией экспрессии РНК. На восьмой неделе в образцах ткани была обнаружена сильная левосторонняя латерализация метилирования CpG островка, обозначающая более сильную репрессию генной транскрипции в левой половине спинного мозга. Это соответствует нашим результатам о повышенной общей правосторонней экспрессии генов на тот момент времени. Прямое сравнение расположения асимметрично метилированных CpG островков и асимметрично экспрессированных генов указывало на то, что 27% вариаций асимметричной экспрессии генов на восьмой неделе можно объяснить посредством дифференциального метилирования. На 10 неделе асимметрия метилирования в образцах ткани значительно снизилась, по сравнению с восьмой неделей, что также соответствует данным об экспрессии генов. Трудно провести интерпретацию на 12 неделе, так как здесь анализировался только один образец, в котором намного повысилось количество асимметрично метилированных CpG сайтов.
С другой стороны, нам также удалось продемонстрировать, что асимметрия при экспрессии генов модулируется посредством асимметрии экспрессии микроРНК. Особенно интересным оказался наш результат, говорящий о том, что для асимметрично экспрессированных микроРНК относительно правой половины спинного мозга на восьмой неделе анализ метаболических путей KEGG выявил значительный эффект сигнального пути TGF-β. Это весьма любопытный результат, так как и центральный фактор роста (Nodal), и лево-правый определяющий фактор (Lefty), два основных белка для создания в теле лево-правой асимметрии в процессе развития, являются частью суперсемейства TGF-β (Миттвох, 2008; Ширатори и Хамада, 2014). Важно отметить, что сигнальный путь TGF-β напрямую связан с леворукостью или праворукостью, так как пропротеинконвертаза 6-го субтилизин-кексинового типа 6 (PCSK6), один из главных кандидатных генов для леворукости или праворукости, кодирует протеазу, которая расщепляет белок NODAL (Скерри и др., 2011).
Наш сбор информации ограничивался неделями с 8 по 12 после зачатия, и для последующих исследований может быть потенциально интересно включить образцы тканей даже с более ранних стадий для получения более подробной картины динамики развития. Также необходимо провести независимую ауторепродукцию более крупных образцов с целью получения углубленных функциональных выводов. В связи с тем, что предпочтения в использовании той или иной конечности были отмечены у многих приматов, отличных от человека, строгое распределение 90 к 10 праворукости и леворукости у людей кажется скорее уникальным, и сравнительный анализ образцов тканей приматов мог бы предоставить уникальные сведения относительно эволюции молекулярной основы межполушарной асимметрии.
Один потенциальный вопрос относительно интерпретации наших данных состоит в том, что объем наблюдаемых асимметрий экспрессии генов связан с висцеральной позицией, и необязательно с асимметрией нервной системы. Особенно интересным экспериментом в этой связи было бы проведение исследования асимметрии экспрессии генов спинного мозга при извращенном положении внутренностей (iv) у мутантов мыши (Окада и др., 1999). Для данных мутантов характерна неупорядоченная латеральность внутренних органов и, при сравнении асимметрии экспрессии генов спинного мозга у животных с нормальными и нетипичными асимметриями внутренностей, можно легко идентифицировать потенциальное воздействие асимметрии внутренностей на экспрессию генов спинного мозга.
В своей совокупности, данные результаты представляют собой сильный индикатор эпигенетического воздействия на асимметрию экспрессии генов спинного мозга у человека, что является потенциальным предшественником леворукости или праворукости. Для птиц было продемонстрировано, что поведенческое предпочтение поворота головы вправо в результате эпигенетической модуляции во время критического периода сразу перед вылупливанием, вызывает не только моторные, но и визуальные, а также когнитивные асимметрии (Кейси и Мартино, 2000; Маннс и Гюнтюркюн, 2009; Роджерс, 1982; Скиба и др., 2002). Похожая модель, основанная на наших данных, применима и к человеку. На восьмой неделе после зачатия появляются скоординированные движения руками, и поведенческие асимметрии рук возникают впервые именно в этот момент (Хеппер и др., 1998; де Врайс и др., 1985), мы предполагаем, что определенные временные рамки до 10 недель после зачатия являются критическим периодом для формирования леворукости или праворукости. В течение данного периода асимметрическое метилирование ДНК и посттранскриптуальная регуляция посредством асимметрично экспрессированными микроРНК приводят к резкому росту числа асимметрий экспрессии РНК в спинномозговых сегментах, иннервирующих руки и кисти. Такие асимметрии экспрессии генов, релевантные для развития центральной нервной системы, могут привести к дифференциальному развитию нейронных цепей, оказывающих влияние на правую руку и кисть, вызывая описанную поведенческую асимметрию. Например, было продемонстрировано, что сегменты спинного мозга, иннервирующие правую руку, содержат мотонейроны с более крупной сомой, чем у сегментов левой руки. Для сравнения, у сегментов левой и правой половины, иннервирующих верхнюю часть торса, нет разницы в размерах (Мельбах и др., 1996). Более того, на восьмой неделе после зачатия спинной мозг и двигательная область коры головного мозга функционально не связаны между собой (тен Донкелар и др., 2004), что также поддерживает предположение о том, что поведенческая асимметрия при использовании рук на этой стадии контролируется спинным мозгом. На более поздней стадии развития, когда спинной мозг и двигательная область коры головного мозга функционально связаны между собой, возникшая поведенческая асимметрия может стать причиной асимметрий в процессах нейрональной пластичности, зависящих от использования (Кирилло и др., 2010), в двигательной области коры головного мозга, которые, в конце концов, приводят к соотношению с леворукостью или праворукостью в коре мозга (Окленбург и др., 2013с). Данный процесс может начаться примерно с 14 недели после зачатия, так как асимметричное использование рук плодом на этой стадии строго соотносится с леворукостью или праворукостью в школьном возрасте (Хеппер и др., 1990, 2005). В отличие от моделей, которые предполагают, что леворукость или праворукость в первую очередь контролируется аллельным разнообразием одного или нескольких кандидатных генов (Аннетт, 1998), мы считаем, наряду с результатом, что более 75% вариаций в сведениях о леворукости или праворукости объясняются факторами окружающей среды, несвязанными с генами (Медланд и др., 2009). Более того, наши данные не противоречат ни анализам групп сцеплений при расширенном изучении источника происхождения леворукости, ни общегеномным ассоциативным исследованиям, которые не были способны идентифицировать аллельные варианты, объясняющие не только фракцию вариаций в сведениях о леворукости или праворукости. (Армур и др., Эрикссон и др., 2010; Сомерс и др., 2015). При этом наши результаты позволяют сделать предположение, что большая часть данных факторов влияния действует в предродовой период, также на формирование актуального доминирования одной из рук воздействуют важные перинатальные и постнатальные факторы окружающей среды, такие как социальная модуляция (Шаафсма и др., 2009).
Подводя итог, следует отметить, что мы смогли продемонстрировать явные, зависимые от времени асимметрии экспрессии генов в тканях спинного мозга у человеческого плода, что связано с зарождением поведенческой асимметрии. Поэтому, согласно нашим данным, можно предположить, что в спинном могзе, а не в коре головного мозга начинаются межполушарные асимметрии. Рассматриваемые асимметрии экспрессии генов модулировались посредством метилирования асимметричного CpG островка и асимметрией при экспрессии микроРНК, наталкивая на вывод, что данные процессы формируют молекулярную основу эпигенетики асимметрии. В заключение, наши данные дают веские основания предполагать о наличии многофакторной модели онтогенеза межполушарных асимметрий, включая множественные как генетические, так и эпигенетические факторы.
Перевод с английского,
Источник
Материал переведен и подготовлен Командой «ПравоНалево»
Перепечатка материала возможна только при указании прямой ссылки на страницу материала и упоминании в тексте материала:
Клуб левшей «ПравоНалево»
Основатель клуба — Елена Боева
propravonalevo.com
Рекомендуемые материалы:
ОТЧЕГО ЖИЗНЬ ТАКАЯ ЛЕВОРУКАЯ? ВИНОВАТЫ ЗВЕЗДЫ
НЕЙРОННЫЕ СВЯЗИ ВАШЕГО МОЗГА РАССКАЖУТ, С КЕМ ВЫ БУДЕТЕ ДРУЖИТЬ!
РУКОВОДИТЕЛЬ — АМБИДЕКСТР. ВЫГОДНО ЛИ БЫТЬ ИМ?
Влияет ли выбор способа вскармливания ребенка на леворукость. Грудное или искусственное?
{kind=link}